Всі еукаріотичні клітини містять набір основних обмежених мембраною органел
Багато важливих біохімічних процесів протікають усередині мембран або на їх поверхнях. Наприклад, при окислювальному фосфоруванні і при фотосинтезі потрібна напівпроникна мембрана для сполучення транспорту протонів з синтезом АТР. Більш того, мембрани служать каркасом для синтезу своїх власних компонентів. Внутрішні мембрани еукаріотичної клітини роблять можливою функціональну спеціалізацію різних мембран, що є, як ми побачимо, вирішальним чинником в розділенні безлічі різних процесів, що протікають в клітині.
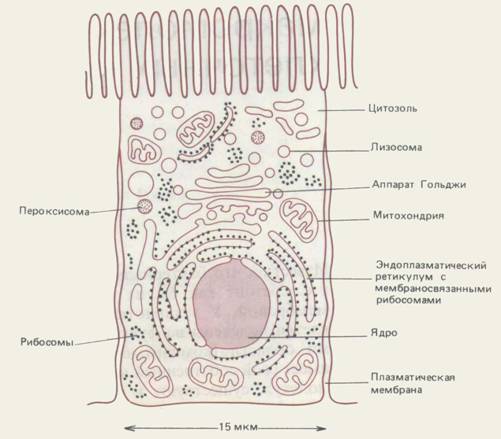
Мал. 8-1. Схематичне зображення основних внутрішньоклітинних компартментів типової тваринної клітини. Цитозоль, ендоплазматичний ретікулум, апарат Гольджі, ядро, мітохондрія, ендосома, лізосома і пероксисома є індивідуальними компартменти, відокремлені від решти клітини принаймні однією вибірковопроникною мембраною.
Внутрішньклітинні компартменти, загальні для всіх еукаріотчних клітин, показані на мал. 8-1. Ядро містить основну частину генома і є головним місцем синтезу ДНК і РНК. Цитоплазма, що оточує ядро, складається з цитозоля і розташованих в ньому цитоплазми органел. Об'єм цитозоля складає трохи більше половини від загального об'єму клітини. Саме у ньому синтезується білок і протікають більшість реакцій так званого проміжного обміну - тобто реакцій, в яких одні малі молекули руйнуються, а інші утворюються, забезпечуючи необхідні будівельні блоки для синтезу макромолекул. Близько половини всіх мембран клітини обмежують схожі на лабіринт порожнини ендоплазматичного ретікулума (ЕР). На зверненій до цитозолю стороні ЕР знаходиться безліч рибосом. Ці рибосоми зайняті синтезом інтегральних мембранних білків і розчинних білків, призначених для секреції або для інших органел. У ЕР також синтезуються ліпіди для решти всієї клітини. Апарат Гольджі складається з правильних стопок ущільнених мембранних мішечків, званих цистернами Гольджі; він отримує з ЕР білки і ліпіди і відправляє ці молекули в різні пункти всередині клітини, попутно піддаючи їх ковалентним модифікаціям. Мітохондрії і хлоропласти рослинних клітин виробляють більшу частину АТР, використовуваного в реакціях біосинтезу, що вимагають надходження вільної енергії. Лізосоми містять травні ферменти, які руйнують відпрацьовані органели, а також частинки і молекули, поглинені клітиною ззовні шляхом ендоцитоза. На шляху до лізосом поглинені молекули і частинки повинні пройти серію органел, званих ендосомами. Нарешті, пероксисоми (відомі також як мікротільця) є маленькими бульбашками, що містять безліч окислювальних ферментів.
Окрім тільки що перерахованих головних мембранних органелл, клітина містить безліч дрібних бульбашок, службовців переносниками речовин між органелламі, а також бульбашок, що зв'язуються з плазматичною мембраною в процесах ендоцитоза і секреції.
В цілому, кожна мембранна органела володіє як властивостями загальними для будь-яких клітин, так і специфічними особливостями, пов'язаними із спеціалізованими функціями диференційованих клітин. Всі разом мембранні органели займають близько половини об'єму клітини. Цілком природно, що для їх побудови потрібна велика кількість внутрішньоклітинних мембран. У двох типах клітин ссавців загальна поверхня мембран ендоплазматичного ретикулума перевищує поверхню плазматичної мембрани відповідно в 25 і в 12 разів. Якщо говорити про масу і площу, то плазматична мембрана в більшості еукаріотичних клітин - всього лише невелика частина всіх мембран.
|
